 Slnečné žiarenie je primárnym zdrojom energie pozemského života. Z fyzikálneho pohľadu je energia širokospektrálneho (teda aj slnečného) žiarenia pohlcovaná tým efektívnejšie, čím širšie rozmedzie vlnových dĺžok fotónov (počnúc časťou spektra, v ktorej zdroj vyžaruje maximum energie) a čím väčšie percento fotónov každej z týchto vlnových dĺžok je zachytávané. Každý nevyužitý fotón by totiž spôsoboval čiastočnú stratu dostupnej energie. Rovnomerné pohlcovanie fotónov všetkých vlnových dĺžok v rozmedzí spektra viditeľného žiarenia (keďže práve v tejto časti elektromagnetického spektra Slnko vďaka svojej povrchovej teplote vyžaruje maximum energie) je ľudským mozgom interpretované ako čierna farba - preto sú čierne napr. aj solárne články. Aplikácia v biológii nám generuje priamočiary záver - ak by mali pozemské rastliny využívať energiu slnečného žiarenia s maximálnou možnou efektivitou jej premeny na chemickú energiu (ktorá je hybnou silou živých organizmov na Zemi), tak z fyzikálno-chemického hľadiska by mali jednoznačne rovnomerne pohlcovať čo najviac fotónov spadajúcich do viditeľnej časti spektra, a to i bez ohľadu na ďalšie procesy nasledujúce po zachytení fotónu. Bezprostredne sa nám tak vynára neobvyklá no logická otázka týkajúca sa jednej zo základných charakteristík pozemskej prírody okolo nás - prečo rastliny nie sú čierne?
Slnečné žiarenie je primárnym zdrojom energie pozemského života. Z fyzikálneho pohľadu je energia širokospektrálneho (teda aj slnečného) žiarenia pohlcovaná tým efektívnejšie, čím širšie rozmedzie vlnových dĺžok fotónov (počnúc časťou spektra, v ktorej zdroj vyžaruje maximum energie) a čím väčšie percento fotónov každej z týchto vlnových dĺžok je zachytávané. Každý nevyužitý fotón by totiž spôsoboval čiastočnú stratu dostupnej energie. Rovnomerné pohlcovanie fotónov všetkých vlnových dĺžok v rozmedzí spektra viditeľného žiarenia (keďže práve v tejto časti elektromagnetického spektra Slnko vďaka svojej povrchovej teplote vyžaruje maximum energie) je ľudským mozgom interpretované ako čierna farba - preto sú čierne napr. aj solárne články. Aplikácia v biológii nám generuje priamočiary záver - ak by mali pozemské rastliny využívať energiu slnečného žiarenia s maximálnou možnou efektivitou jej premeny na chemickú energiu (ktorá je hybnou silou živých organizmov na Zemi), tak z fyzikálno-chemického hľadiska by mali jednoznačne rovnomerne pohlcovať čo najviac fotónov spadajúcich do viditeľnej časti spektra, a to i bez ohľadu na ďalšie procesy nasledujúce po zachytení fotónu. Bezprostredne sa nám tak vynára neobvyklá no logická otázka týkajúca sa jednej zo základných charakteristík pozemskej prírody okolo nás - prečo rastliny nie sú čierne?
Úvod
Tento článok je určený všetkým čitateľom, ktorí radi premýšľajú nad netradičnými otázkami a majú potešenie z hľadania súvislostí medzi existujúcimi faktami a informáciami, častokrát interdisciplinárnymi. Komplexne sumarizuje a dáva do súvislostí súčasný stav znalostí vedcov o uvedenej téme (pričom čitateľovi zároveň predstaví niekoľko organizmov s nezvyčajnými schopnosťami), a to formou zrozumiteľnou sčítanému laikovi disponujúcemu iba základnými znalosťami z fyziky, chémie a biológie. Rozsah a hĺbka článku bola zároveň zvolená tak, aby bolo v čo najväčšej miere možné vyhnúť sa nepresným zjednodušeniam hroziacim z jeho populárno-vedeckého účelu. Jeho vznik bol motivovaný jednak absenciou ucelených, komplexných a dostatočne racionálne podložených informácií v dostupných zdrojoch, a jednak výsledkami ankety, v ktorej väčšina účastníkov vyjadrila presvedčenie, že najefektívnejšou formou existencie rastlín je farba zelená a nie čierna. Samotná striktne logicky a fyzikálne podložená odpoveď, že tento názor je mylný, je však nedostatočná - je totiž súčasne potrebné i vysvetliť, prečo nám pozemská evolúcia čiernu vegetáciu evidentne neponúka. Poďme sa teda pozrieť zdanlivému nerešpektovaniu fyziky biológiou podrobnejšie na koreň.
Neživá príroda - maximálna efektivita
V neživej prírode majú základné deje jeden spoločný menovateľ - vo všeobecnosti prebiehajú tak, aby nastala minimalizácia energie. Tento princíp je prirodzenou vlastnosťou prírody, nezávislou od pozorovateľa. Dôsledky sa prejavujú najrôznejšími spôsobmi, spomedzi ktorých si uveďme aspoň približne guľový tvar hviezd či kvapiek vody, zodpovedajúci minimálnej potenciálnej energii (samozrejme pri zahrnutí sekundárnych javov ako napr. rotácia hviezd či odpor vzduchu padajúcej kvapky je najefektívnejším tvarom elipsoid či iná deformácia, spĺňajúca však stále podmienku minimalizácie energie). Rovnako sa týmto princípom riadi i pohyb či dráha objektov (napr. obeh planét okolo Slnka, hviezd v galaxiách, balistická krivka jadrovej strely, pohyb pružín v mechanických hodinkách a pod.). Táto charakteristická črta prírody sa vo všeobecnosti vzťahuje dokonca i na kvantové javy, a to napriek mnohým zásadným odlišnostiam medzi kvantovým svetom a makrosvetom (pre matematicky podkutejšieho čitateľa sa zmieňme, že systémy makro- aj mikrosveta vychádzajú z tzv. hamiltoniánu, resp. lagrangiánu, pričom správanie sa systému popisuje minimalizácia, resp. maximalizácia týchto funkcií).
Laickým jazykom sformulované, jedným z experimentálne i teoreticky veľmi dobre doložených základov správania sa prírody je teda minimalizácia či presnejšie optimalizácia energie, inými slovami energetická vyváženosť či efektivita.
Živá príroda - stále v procese evolúcie
V živej prírode je situácia komplikovanejšia. Živá príroda síce procesy s maximálnou efektivitou vytvára, avšak nie vždy a nie hneď. Pri analýze tohto problému si v prvom rade musíme uvedomiť jednu dôležitú skutočnosť - živá príroda je komplexnou nadstavbou prírody neživej, pričom musí postupne optimalizovať množstvo spočiatku aj veľmi neefektívne nastavených vstupných parametrov, používajúc deje a procesy založené na striktných fyzikálnych zákonoch. Tvar hviezd je daný len niekoľkými parametrami, pri komplexných biochemických dejoch ich ale existuje väčší počet. Počiatočný stav zďaleka nemusí byť vhodný k optimalizácii konečného procesu, a vo väčšine prípadov nie je možné vylepšiť existujúci stav kompletným 'revolučným' prebudovaním celého komplexného procesu (pretože globálna optimalizácia si kvôli zachovaniu funkčnosti celku vyžaduje súčasnú zmenu celej série navzájom súvisiacich parametrov, ktorá môže zo štatistického hľadiska nastať len s veľmi malou pravdepodobnosťou). Živá príroda je teda odkázaná na postupnú a zdĺhavú optimalizáciu jednotlivých parametrov (navyše ešte v neustále sa meniacich podmienkach), ktoré nie vždy vedú z globálneho pohľadu k najefektívnejšiemu a už vonkoncom nie rýchlemu riešeniu.
Uveďme si však na ilustráciu jeden názorný príklad, kedy aj živá príroda vytvorila v podstate maximálne efektívny proces - ide o tzv. transláciu DNA, pri ktorej je postupnosť nukleotidov nášho genetického kódu (cytozín, guanín, adenín a tymín) 'prekladaná' do konkrétnych bielkovín. Tento proces prebieha tak, že z kópie DNA (tzv. mRNA - messenger RNA), presunutej z jadra do špecializovaných organel bunky (tzv. ribozómov), sa vytvára poradie aminokyselín, ktoré sú základným stavebným prvkom všetkých bielkovín. Preklad genetickej informácie sa uskutočňuje prostredníctvom sady tzv. tRNA (transfer RNA) molekúl, pričom každá z týchto molekúl na jednej svojej strane obsahuje postupnosť nukleotidov a na strane druhej presne definovanú zodpovedajúcu aminokyselinu. Slúži teda ako pevne definovaný „molekulárny slovník“, priraďujúci ku konkrétnej kombinácii nukleotidov konkrétnu aminokyselinu. Do ribozómov teda vchádza kópia DNA a v ňou definovanom poradí postupne molekuly tRNA prinášajúce aminokyseliny, ktoré ribozóm spojí do súvislého reťazca. Poradie aminokyselín následne definuje presný tvar a funkciu bielkovín či enzýmov. Takto sa informácia zakódovaná v DNA pretransformuje na reálnu bielkovinu. Aminokyselín zakódovaných v DNA, z ktorých sa vytvárajú všetky ďalšie aminokyseliny a proteíny, nie je veľa - je ich rovných dvadsať. Aby sme vedeli zakódovať tento počet (ktorý je nevyhnutný pre vybudovanie existujúcich bunkových štruktúr) do DNA, potrebujeme nájsť najmenší počet po sebe nasledujúcich nukleotidov, ktoré môžu zakódovať dostatočný počet aminokyselín, ale zároveň musíme minimalizovať frekvenciu mutácií (čiže možných porúch v poradí nukleotidov a z nich vyplývajúcich chýb) a takisto energetické a surovinové náklady na celý proces kódovania a dekódovania. Ideálny počet z matematického hľadiska sa rovná trom nukleotidom kódujúcim jednu aminokyselinu - toto riešenie problému kedysi navrhol renomovaný teoretický fyzik George Gamow (čítate správne - nie biológ ani biochemik, ale teoretický/matematický fyzik). A čo urobila živá príroda? Evolúcia skutočne zabezpečila najefektívnejšie riešenie - použitie trojíc nukleotidov. Pred vyvinutím vysoko sofistikovaného translačného mechanizmu a pri vyššej prírodnej radiácii (ktorá na Zemi v dávnej minulosti vďaka väčšiemu obsahu uránu a tória v zemskej kôre panovala) pravdepodobne pôvodne používala kódovanie aminokyselín štvoricami nukleotidov (a nie trojicami), a i keď táto možnosť nie je jednoznačne potvrdená, faktom je, že dnes používané trojice sú maximálne efektívnym riešením. Pozoruhodný príklad súladu medzi matematickým riešením a reálnou optimalizáciou v živej prírode na molekulárnej úrovni.
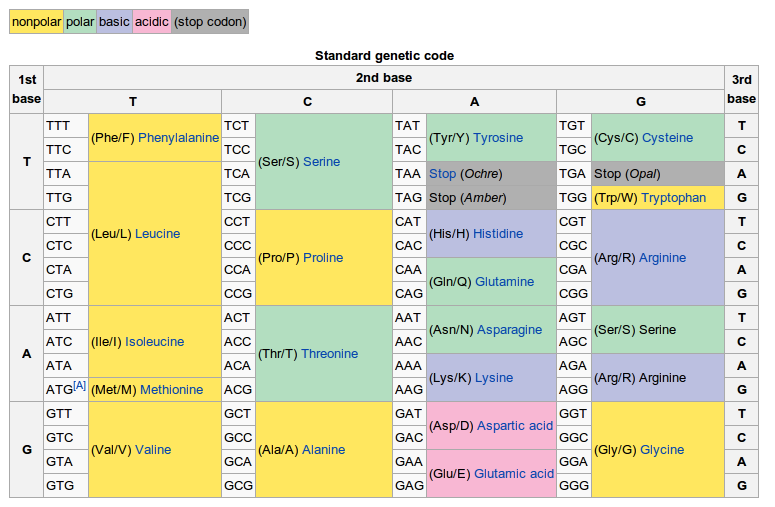
Obr. 1: Názorný príklad procesu, ktorého jednu časť evolúcia stihla z matematického pohľadu maximálne zefektívniť - translácia DNA (resp. mRNA) na proteíny sa uskutočňuje tak, že každá z 20 základných aminokyselín (stavebných kameňov všetkých proteínov) je kódovaná trojicou nukleotidov. Nie je vylúčené, že kedysi príroda používala kvôli zvýšenej rádioaktivite a teda i frekvencii mutácií bezpečnejšie no energeticky a surovinovo menej výhodné kódovanie pomocou štvoríc. Tabuľka obsahuje trojice nukleotidov (tzv. kodóny) kódujúce jednotlivé aminokyseliny, pričom uvedené párovanie je realizované prostredníctvom tRNA molekúl. I keď existencia trojíc je maximálne efektívnym riešením, samotné priradenie kodónov k aminokyselinám je naďalej predmetom evolučného vývoja - jedným z viacerých dôkazov je napr. dôležitá súčasť dnešných buniek, tzv. mitochondria, slúžiaca ako 'energetická továreň' konvertujúca glukózu na bunkou priamo použiteľnú energetickú molekulu ATP. Mitochondria používa mierne odlišné kodóny, čo je s najväčšou pravdepodobnosťou dôsledkom toho, že vznikla - podobne ako chloroplast v rastlinných bunkách - symbiotickým (čiže "spolunažívacím") pohltením dávnych baktérií a takmer bez výnimky si dodnes ponechala časť svojej pôvodnej DNA. Zdroj: wikimedia.
Avšak nie vždy evolúcia prírody mala do dnešného dňa dostatok času optimalizovať procesy až k ich najefektívnejším formám. Jedným z nich môže byť aj fotosyntéza.
Fotosyntéza
Proces, pri ktorom prostredníctvom špecializovaných molekúl dochádza k premene energie fotónov (čiže elektromagnetického žiarenia) na energiu chemickú, ktorá je ďalej využívaná bunkou na realizovanie zložitých biochemických procesov, sa nazýva fototrofia (nemýliť si s odlišným pojmom fototropia). Evolúcia schopnosti fototrofie je veľmi zaujímavá, predovšetkým vďaka tomu, že podľa súčasných znalostí sa s najväčšou pravdepodobnosťou vyvinula iba dvakrát - najprv v skupine evolučne veľmi starých jednobunkových organizmov Archaea (odlišných od baktérií aj ďalších organizmov) využívajúcich molekulu tzv. retinalu (nešlo ešte o fotosyntézu), a po druhý krát v skupine kyanobaktérií využívajúcich molekuly chlorofylu, z ktorých neskôr evolučne (s najväčšou pravdepodobnosťou symbiotickým pohltením) vznikli chloroplasty v dnešných (nielen) rastlinných bunkách. Fototrofia využívajúca skupinu molekúl chlorofylu (resp. veľmi podobného baktériochlorofylu u niektorých baktérií) je nazývaná fotosyntézou.
Retinal je citlivý najmä na zelenú časť spektra, v ktorej na povrch Zeme zo Slnka dopadá veľká časť celkovej energie. Výhodou biosystémov obsahujúcich evolučne mladší chlorofyl či baktériochlorofyl je však predovšetkým možnosť zapojiť do procesu konverzie energie fotónov aj sekundárne pomocné pigmenty, ktoré rozširujú pohltené spektrum, ďalej výroba energeticky bohatšej molekuly (konkrétne sacharidov), ale predovšetkým dôležitá schopnosť zabudovávania anorganickej látky - oxidu uhličitého - do zložitejších organických molekúl, inými slovami transformácia či pretváranie neživej hmoty na hmotu živú. To s retinalom nie je možné. Zapojenie pomocných pigmentov je realizované prostredníctvom vzájomne prepojených tzv. anténnych systémov, ktoré obsahujú zväčša až niekoľko stoviek molekúl fotopigmentov. Ich úlohou je prenášať energiu zo zachytených fotónov do tzv. reakčného centra, kde sa energia koncentruje v centrálnej molekule chlorofylu - pracujú teda skutočne ako "antény" zosilňujúce signál, resp. príjem energie. Reakčné centrá sa už na rozdiel od chlorofylu evolučne vyvinuli postupne pri viacerých udalostiach.
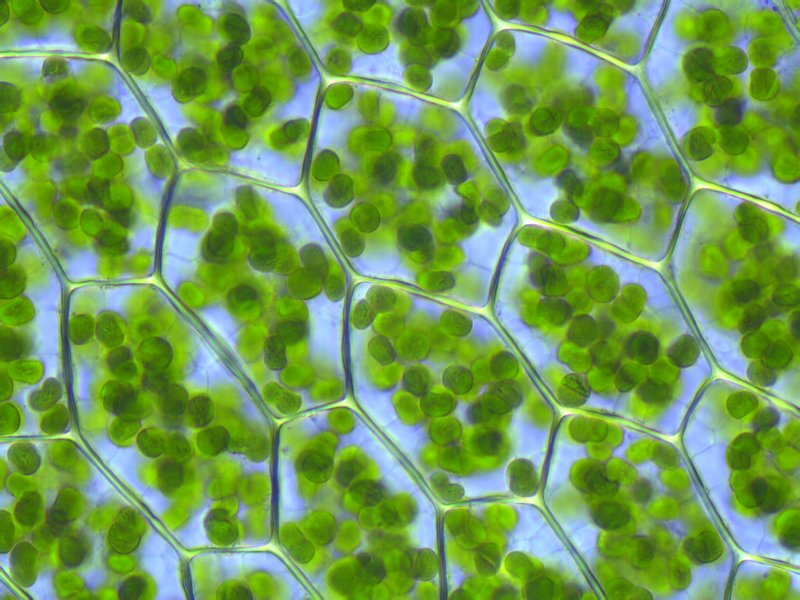 Obr. 2: Chloroplasty v bunkách machu obsahujúce chlorofyl a sekundárne pigmenty zabudované do tzv. anténnych systémov, slúžiacich na rozšírenie absorpčnej schopnosti chlorofylu. Existuje veľké množstvo vedeckých dôkazov na makro aj mikroskopickej (resp. genetickej) úrovni, že chloroplasty vznikli pohltením (tzv. endosymbiózou) dávnych kyanobaktérií. Dodnes sa v nich nachádza časť ich pôvodnej DNA, väčšina génov kódujúcich proteíny nevyhnutné na správnu činnosť chloroplastov sa však už presunula do jadra bunky. Zdroj: wikimedia.
Obr. 2: Chloroplasty v bunkách machu obsahujúce chlorofyl a sekundárne pigmenty zabudované do tzv. anténnych systémov, slúžiacich na rozšírenie absorpčnej schopnosti chlorofylu. Existuje veľké množstvo vedeckých dôkazov na makro aj mikroskopickej (resp. genetickej) úrovni, že chloroplasty vznikli pohltením (tzv. endosymbiózou) dávnych kyanobaktérií. Dodnes sa v nich nachádza časť ich pôvodnej DNA, väčšina génov kódujúcich proteíny nevyhnutné na správnu činnosť chloroplastov sa však už presunula do jadra bunky. Zdroj: wikimedia.
Existujú dva základné typy fotosyntézy – prvým je tzv. anoxygénna fotosyntéza, pri ktorej nedochádza k produkcii kyslíka. Druhým je fotosyntéza oxygénna, pri ktorej je kyslík produkovaný ako vedľajší produkt, a ktorá je – a to je podstatné - v porovnaní s anoxygénnou fotosyntézou energeticky efektívnejšia, t.j. z rovnakého množstva energie fotónov vytvorí viac chemickej energie. Ako sa môžeme ľahko presvedčiť jednoduchým nádychom, príroda evolučne jednoznačne zvýhodnila a naďalej preferuje energeticky výhodnejšie riešenie procesu produkujúce kyslík. Anoxygénnu fotosyntézu uskutočňujú dnes len niektoré druhy extremofilných baktérií (napr. termofilných, t.j. žijúcich v horúcich sopečných prameňoch, kde rastliny a iné oxygénne fotosyntetizujúce organizmy nedokážu prežiť), zatiaľ čo rastliny, riasy a väčšina ostatných fotosyntetizujúcich baktérií uskutočňujú výhodnejšiu oxygénnu formu fotosyntézy. Pri tomto dominantnom type je - zjednodušene povedané - energia fotónu použitá na rozštiepenie molekuly vody (voda je teda na rozdiel od anoxygénnej fotosyntézy nevyhnutná), kyslík uniká ako vedľajší produkt a vodík sa zlučuje s oxidom uhličitým za vzniku na chemickú energiu bohatej molekuly. Samozrejme to všetko prebieha prostredníctvom sofistikovaného komplexu biomolekúl, chemických reakcií a fyzikálnych dejov (napr. prenosu elektrónov v dvoch rôznych chlorofylových fotosystémoch, či dvoch fáz fotosyntézy – jednej závislej od svetla, druhej nezávislej), ich detaily však pre tento článok nie sú relevantné (tým je len výsledná účinnosť týchto systémov), takže sa ňou v záujme zachovania relatívnej stručnosti nebudeme podrobnejšie zaoberať.
Presuňme sa však zo všeobecnej roviny ku konkrétnejším faktom. Všetky dnešné organizmy teda obsahujú len neveľkú škálu fotopigmentov, z ktorých primárny je chlorofyl (presnejšie viacero jeho typov, z ktorých najdôležitejšie sú 'chlorofyl a' a 'chlorofyl b'), pričom zmieňovanými pomocnými fotopigmentmi sú karotény dávajúce napr. aj mrkve jej oranžovú farbu, a xantofyly sfarbujúce listy stromov na jeseň do žlta. Chlorofyl dokáže pohltiť veľkú časť fotónov v modrej a červenej oblasti spektra, no má takmer nulovú absorpciu fotónov v zelenej oblasti. Avšak s pomocou anténnych systémov obsahujúcich okrem chlorofylu i zmienené sekundárne fotopigmenty je už u rastlín situácia výrazne lepšia - pre väčšinu vlnových dĺžok viditeľného žiarenia dokáže komplex fotopigmentov pohltiť viac než 90% energie žiarenia, čo je bezpochyby obdivuhodná efektivita. Napriek tomu žiaden z týchto pigmentov nedokáže efektívne absorbovať fotóny s energiou zodpovedajúcou zelenej farbe, presnejšie povedané aj ich kombináciou vznikne situácia, že zelená časť spektra sa pohlcuje s menšou účinnosťou než ostatné energie. Pre upresnenie - neznamená to, žeby rastliny túto oblasť spektra vôbec neboli schopné pohltiť, naopak, s pomocou sekundárnych pigmentov list priemernej rastliny dokáže pohltiť viac než polovicu dopadajúcich fotónov zelenej farby - čo je však málo oproti viac než 90 percentnej účinnosti pri ostatných fotónoch. Pritom práve v zelenej časti spektra povrch Zeme a vrchná časť oceánov dostáva významnú časť prísunu energie elektromagnetickým žiarením.
Situácia sa týka rovnako aj mnohých dnešných fotosyntetizujúcich baktérií (ktoré vznikli zo spoločného evolučného predka ako rastlinné chloroplasty) a analogicky dokonca i zelených rias, z ktorých žiadne nedokážu pohlcovať rovnomerne celú oblasť spektra.
Čiastočnú výnimku však tvoria niektoré špecifické druhy fytosyntetizujúcich baktérií a nezelených rias, ktoré sa k tomuto cieľu dopracovali bližšie než bežné rastliny. Dosiahli to vývojom ďalšej skupiny sekundárnych fotopigmentov, tzv. fykobilínov (k nim sa ešte v nasledujúcej kapitole vrátime), vďaka čomu sa javia byť čierne, avšak je to len ilúzia spôsobená kontrastom s intenzitou okolitého osvetlenia, v skutočnosti ani oni nepohlcujú rovnomerne celú oblasť viditeľného spektra.
Pre úplnosť sa zmieňme i o skupine machorastov odolných voči nízkym teplotám, ktoré neobsahujú fykobilíny a napriek tomu sa javia byť čierne - i oni sú však v skutočnosti iba tmavo zelené.
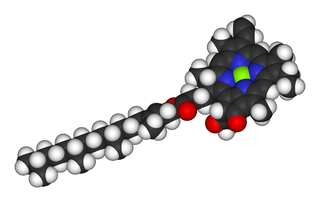
Obr. 3: Chlorofyl je funkčne vysoko sofistikovanou molekulou, ktorej evolučný vznik si vyžiadal nezanedbateľne dlhý čas. Existuje viacero jeho druhov, ktoré sa však líšia len detailmi svojej štruktúry. Na obrázku vidíme konkrétne ‚chlorofyl a’. Anoxygénne fotosyntetizujúce baktérie používajú ďalšiu skupinu príbuzných molekúl tzv. baktériochlorofylov. Štrukturálna a funkčná podobnosť všetkých týchto fotopigmentov je indíciou, že sa vyvinuli z molekuly, ktorá vznikla pri jednorazovej evolučnej udalosti. Zdroj: wikimedia.
 N
N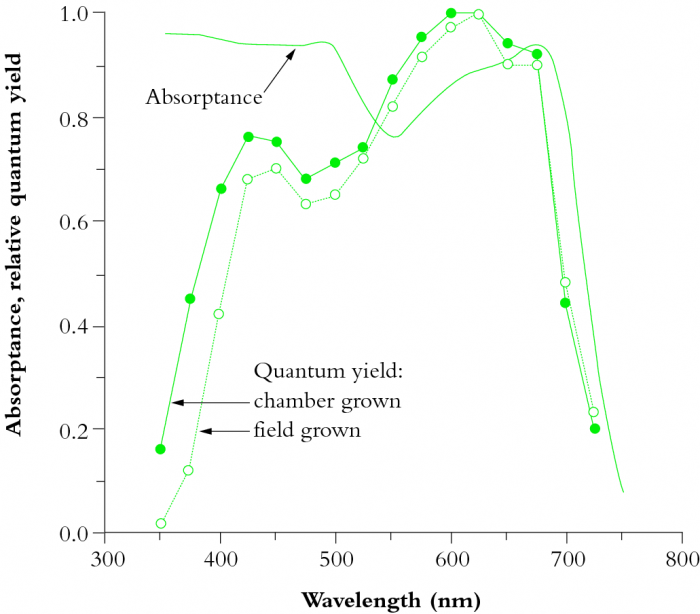
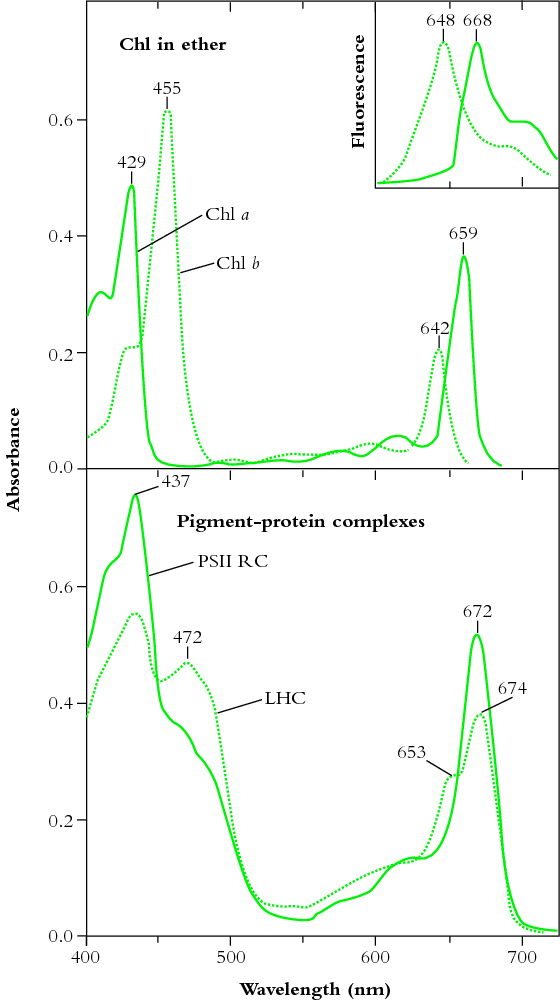
Obr. 4: Napriek tomu, že celková efektivita chlorofylu a využitie s ním spojených proteínových komplexov sú obdivuhodné (a výrazne vyššie než u jednoduchšieho retinalu), nie je ideálna. V ľavej hornej časti obrázku je znázornená absorpčná schopnosť izolovaných molekúl dvoch základných typov chlorofylu a (Chl a) a chlorofylu b (Chl b), v ľavej spodnej časti vidíme absorpčnú schopnosť týchto chlorofylov zabudovaných do štandardných pigment-proteínových komplexov (tzv. reakčných centier), ktoré sa vyskytujú v prírode. Ako vidíme, zelená časť spektra zodpovedajúca vlnovej dĺžke cca 500-570 nm nie je čistým chlorofylom takmer vôbec pohlcovaná, a aj jeho účinnosť v reakčných centrách (ktorá zodpovedá reálnemu stavu v prírode) pre túto oblasť spektra vykazuje len mierne zlepšenie. V pravej časti obrázku vidíme celkovú absorpčnú schopnosť týchto komplexov aj s využitím všetkých sekundárnych pigmentov (znázornenú krivkou bez krúžkov/bodov). Tam už je situácia pozitívnejšia - aj v zelenej časti spektra chlorofylové komplexy dokážu pohltiť viac než polovicu fotónov. Pre komplexnosť obrázok obsahuje aj dve krivky s plnými a prázdnymi krúžkami - znázorňujú tzv. 'quantum yield', ktorý zodpovedá schopnosti buniek fotón nielen pohltiť, ale i priamo využiť na produkciu energetických molekúl. Plné krúžky zodpovedajú rastlinám vypestovaným pod umelým osvetlením, zatiaľ čo prázdne krúžky zodpovedajú údajom nameraným pri rastlinách vypestovaných v prirodzených vonkajších podmienkach. Rozdiel je daný tým, že vo vonkajšom prostredí pestované rastliny produkujú viac špecializovaných molekúl brániacich radiačnému poškodeniu buniek UV žiarením, čo má však za následok, že rastlina pestovaná vo vonkajšom prostredí dokáže modrú časť spektra na produkciu sacharidov využiť s menšou účinnosťou. Za pozornosť však stojí celkový nízky quantum yield v modrej časti spektra (a to i pre skleníkové rastliny) - ten je hlavným dôvodom problémov s prehrievaním chlorofylových komplexov a ešte sa k nemu v článku vrátime. V každom prípade však vidíme, že aj napriek evidentnej a čiastočne úspešnej snahe prírody kompenzovať nedostatky chlorofylu s pomocou anténnych systémov, má chlorofylový komplex v zelenej časti viditeľného spektra stále najhoršiu efektivitu. Zdroj: Australian Society of Plant Scientists, New Zealand Society of Plant Biologists, and New Zealand Institute of Agricultural and Horticultural Science (Plants in Action).


Obr. 5: Vďaka schopnosti čiastočne absorbovať UV žiarenie sa niektoré fotopigmenty môžu zúčastňovať na ochrane buniek pred UV žiarením, napr. tzv. mutatoxantín v lišajníku Xanthoria elegans mu nielenže dáva oranžovo-žlté sfarbenie, ale je jednou z príčin, prečo tento lišajník dokáže prežiť aj dlhé obdobie v nechránenom vesmíre (Xantoria elegans prežil v otvorenom a pred vákuom ani radiáciou nechránenom priestore jeden a pol roka počas orbitálneho experimentu na ISS, a je to jeden z najvhodnejších kyslík produkujúcich organizmov na prípadné terraformovanie iných planét). Zdroj: Per Verdonk (vľavo), The University of British Columbia (vpravo).
Prečo živá príroda nie je maximálne efektívna?
Po ozrejmení pozadia a súvislostí sa teda dostávame k pôvodnej otázke - prečo rastliny nevyužívajú energiu slnečných fotónov s maximálnou možnou efektivitou? Skúsme si zhrnúť fakty, ktoré sú pre získanie odpovede relevantné:
A. Ako sme si ukázali, fotosyntéza je závislá na konkrétnych fotopigmentoch, ktoré kvôli fyzikálnym vlastnostiam atómov a molekúl nie sú rovnomerne citlivé na všetky vlnové dĺžky fotónov. Vyvinúť molekulu, ktorá by bola citlivá rovnomerne na fotóny širokého spektra energií, a ktorá by súčasne dokázala byť zapojená do komplexného cyklu premeny energie na bunkou ďalej využiteľné energeticky bohaté molekuly, nie je jednoduché (a iba o niečo menej náročné je vyvinúť funkčný širokospektrálny anténny systém). Evolúcia prírody nie je ešte ukončená, a teda je s vysokou pravdepodobnosťou ešte možné vyvinúť efektívnejšie do fotosystémov zabudovateľné fotopigmenty či ich kombináciu, ktorá bude pokrývať viditeľné spektrum rovnomernejšie. Na to je však potrebný čas, tak ako bol potrebný napr. na vyvinutie spomínaného kódovania proteínov pomocou trojíc bázových párov v DNA, či na vývoj chlorofylových komplexov. Na 'obranu' prírody však opäť musíme zopakovať, že dnešné riešenie je obdivuhodne dobré - z časti spektra, v ktorej na Zem dopadá najviac energie, využíva veľkú väčšinu. Situácia sa dá prirovnať ku kritike dnešného automobilu - áno, existujú aj energeticky efektívnejšie a šetrnejšie riešenia, avšak situácia je výrazne lepšia ako pri prvotných predvojnových spaľovacích automobiloch s vysokou spotrebou paliva. Navyše, príroda už fotopigmenty absorbujúce prevažne zelenú časť spektra vyvinula. Prvou skupinou sú tzv. antokyany, zodpovedné za červenú farbu jabĺk a iných plodov, ktoré však nie je možné zabudovať a využiť v chlorofylových anténnych systémoch, a teda ani priamo zapojiť do fotosyntézy. Druhou skupinou sú už raz zmieňované fykobilíny, ktoré vytvárajú osobitnú formu anténnych systémov (odlišnú od ostatných sekundárnych fotopigmentov), z ktorých niektoré majú absorpčné maximum práve v zelenej časti spektra (predovšetkým fykoerytrín). Výsledný komplex však stále nezabezpečuje rovnomerné pohlcovanie celého viditeľného žiarenia, takže riasy či baktérie, ktoré fykobilíny obsahujú, majú v skutočnosti tmavočervenú alebo tmavo modrozelenú farbu.
Z tohto pohľadu dospela príroda len k čiastočne optimalizovanému bodu, ktorý sa v matematicko-fyzikálnej terminológii nazýva lokálne minimum - niečo, čo je zoptimalizované len vzhľadom na obmedzený počet voľných parametrov. Lokálne minimum však nemusí byť zhodné s globálnym minimom - v tomto prípade s maximálnym pohlteným počtom fotónov v každej časti viditeľného spektra. Je to podobná situácia ako s tekutou vodou v prírode - molekuly vody sa na Zemi snažia minimalizovať svoju potenciálnu energiu tým, že sa premiestňujú do nižšie položených priestorov. Avšak to neznamená, že voda z horského prameňa hneď klesne na úroveň hladiny oceánov - proces poklesu je pozvoľný, vyžaduje si nenulový čas, a môže sa dokonca stať, že načas ustane vďaka prekážke - napr. sa môže vytvoriť jazero pred terénnou vyvýšeninou. V prípade evolúcie je analógiou prekážky nutnosť vyvinúť novú molekulu či zahrnúť do existujúceho procesu nové molekuly. Je možné, že takúto molekulu či molekulárny komplex evolúcia vyvinie v priebehu nasledujúcich stoviek miliónov rokov, a je možné, že sa jej túto bariéru nepodarí prekonať nikdy.
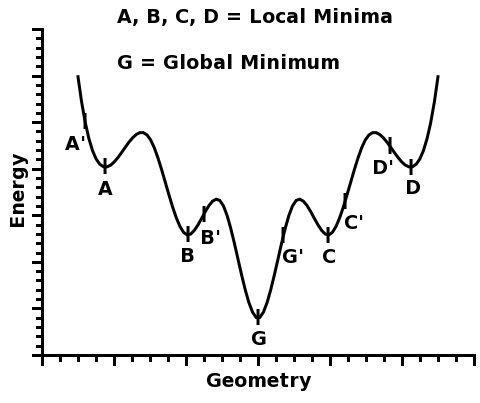
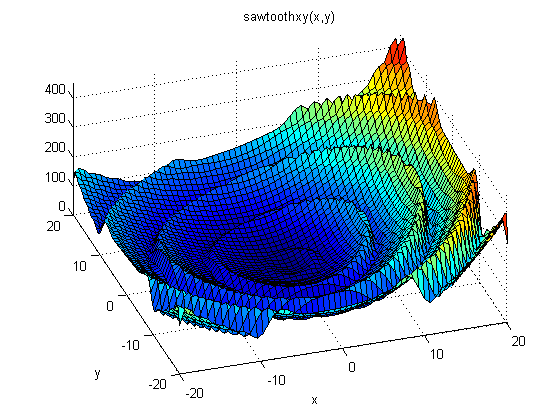
Obr. A-1: Lokálne a globálne minimum funkcie či deja. V ľavej časti obrázku vidíme príklad optimalizácie geometrického tvaru molekuly voči jej energii danej fyzikálnymi a chemickými zákonmi (analogickej k snahe kvapky vody získať guľový tvar). Vidíme, že v závislosti od počiatočného stavu, resp. tvaru molekuly (zodpovedajúcej na obrázku bodom označeným písmenami s čiarkou) môžeme skončiť v jednom z len čiastočne optimalizovaných stavov (body označené písmenami A až D), v ktorých už nie je možné ďalej znižovať energiu molekuly bez kvalitatívnej zmeny a dostať sa až ku globálnemu minimu (na obrázku označenom bodom G). V prípade molekuly je touto kvalitatívnou zmenou dodanie dodatočnej energie, v prípade optimalizácie fotosyntézy v evolúcii života je to vyvinutie novej molekuly či anténneho systému. V strednej časti obrázku vidíme matematický model analogickej situácie pri súčasnej optimalizácii troch parametrov - vytvorí sa tým trojrozmerná optimalizačná plocha s "údoliami" a bariérami. Situácia v prírode môže zahŕňať ešte viac parametrov so zložitejšou viacrozmernou optimalizačnou plochou, preto je existencia lokálnych miním, v ktorých počas vývoja príroda "uviazne", veľmi pravdepodobná, častá a dá sa povedať, že takmer nevyhnutná. Princíp správania sa prírody v zmysle minimalizácie energie prevzala biológia do svojho popisu dejov a evolúcie vo forme tzv."fitness landscape" (voľne preložiteľné napr. ako "plocha adaptivity"), ktorej ilustráciu vidíme na pravom obrázku (za zjednodušujúceho stavu, ak by genetická výbava bola iba dvojrozmernou veličinou).
Zdroje: vľavo - Lanthanide Complexes Computational Chemistry RM1 Research; v strede - MathWorks, Inc.; vpravo - Yale University.

Obr. A-2: Evolúcia sa neskončila, je to neustále prebiehajúci proces. Na obrázku vidíme názornú ukážku pokračujúcej evolúcie - morského slimáka druhu Elysia chlorotica. Ak vám jeho farba nápadne pripomína farbu rastlín, nemýlite sa - je to jeden z mála zástupcov ríše živočíchov, ktorý dokáže priamo uskutočňovať fotosyntézu. Štruktúra a zloženie živočíšnej bunky sa od rastlinnej bunky značne líši, medzi iným aj tým, že živočíšna bunka samozrejme neobsahuje chloroplasty. Živočích Elysia chlorotica však vo svojich bunkách chloroplasty má. Nedokáže si ich zatiaľ vytvárať sám, získava ich pohltením rias. V každom prípade však boli vedci prekvapení, keď zistili, že chloroplasty sú zakomponované skutočne priamo do vnútra buniek slimáka (a nie medzi ne), a keď genetická analýza ukázala, že slimák už má vo svojej DNA gény na ich podporu, vďaka ktorým dokáže chloroplastom zabezpečiť dlhodobú (i keď nie neobmedzenú) stabilitu a existenciu. Vydrží teda dlhé mesiace bez potravy, pretože fotosyntézou vo vlastných bunkách získava cukry a uhlík potrebný pre život. Sme tu svedkami podobnej udalosti (i keď zatiaľ len v medzištádiu) aká nastala pred miliardami rokmi, keď fotosyntetizujúce baktérie boli endosymbioticky pohltené inými bunkami. V budúcnosti bude možno fotosyntézou disponovať omnoho širšia skupina organizmov ako dnes, avšak ako vidíme, stále sa schopnosť fotosyntézy odvíja od jedinej evolučnej udalosti vzniku molekuly chlorofylu. Aj to sa však môže v budúcnosti zmeniť. Zdroj: Patrick Krug, California State University.



Obr. A-3: Rastlinné chloroplasty sa s najväčšou pravdepodobnosťou vyvinuli pohltením predkov dnešných kyanobaktérií (známych aj pod názvom sinice), ktoré ako prvé vyvinuli aj samotný chlorofyl. Keďže chlorofyl po pohltení fotónu používa na svoju neutralizáciu štiepenie vody na kyslík, protón a elektrón, ako odpad sa pri jeho použití uvoľňuje kyslík. Vďaka tejto efektívnejšej oxygénnej fotosyntéze to boli práve kyanobaktérie, ktoré pred miliardami rokov zapríčinili okysličenie zemskej atmosféry a tým aj jedno z najväčších vymieraní v histórii života na Zemi (tzv. kyslíková katastrofa, pri ktorej vyhynuli takmer všetky vtedajšie formy života – pre podrobnosti viď napr. článok o histórii pozemskej klímy a vyhynutí mamutov). Dodnes si táto revolučná skupina baktérií zachovala status jedného z najprogresívnejších a najvynaliezavejších fotosyntetizujúcich organizmov, a v priebehu miliárd rokov rozhodne nezaháľala. Vďaka vyvinutiu špecializovaných fotopigmentov (fykoerytrínov) sú niektoré druhy kyanobaktérií schopné účinne pohlcovať aj zelenú časť spektra. To niektorým z nich dokonca umožňuje na rozdiel od rastlín flexibilne meniť vzájomný pomer fotopigmentov podľa aktuálnych svetelných podmienok, a teda do značnej miery optimalizovať fotosyntézu a zvyšovať jej efektivitu. Rovnomerné pokrytie celého viditeľného spektra, a teda maximálnu efektivitu a čiernu farbu, tým však ani ony nikdy nedosahujú. V každom prípade, vďaka schopnosti fykoerytrínu pohlcovať prevažne zelenú časť spektra je efektivita fotosyntézy u týchto baktérií vo všeobecnosti vyššia než u iných fotosyntetizujúcich organizmov. Je to jeden z dôvodov, prečo sú tieto baktérie evolučne extrémne úspešné a v prírode veľmi rozšírené (ďalšími príčinami sú napr. schopnosť fixovať dusík z atmosféry či existovať i v prostredí bez kyslíka). Globálne na celej Zemi sú kyanobaktérie zodpovedné za cca 20 až 30 percent planetárnej fotosyntézy a na povrchu oceánov uskutočňujú dokonca až 50 percent celkovej fotosyntézy planktónu. To zodpovedá konverzii slnečného žiarenia na chemickú energiu na úrovni približne 450 TW (terawattov; pre porovnanie, celková spotreba energie ľudstvom je cca 15 TW). Kyanobaktérie nájdeme takmer v každom prostredí - v grónskom ľadovci, antarktických jazerách, pustých skalách, vnútri hornín, púšťach, v hĺbkach morí, v extrémne horúcich (až 73 °C), slaných (Mŕtve more) i zásaditých (až pH=13,5, rekord pre akýkoľvek organizmus) prostrediach, a dokonca i vnútri mnohých iných organizmov. Skúsenosť s nimi majú okrem iného aj majitelia bazénov či umelých nádrží, kde sa zdanlivo „čierne“ kyanobaktérie (z ktorých niektoré druhy produkujú nebezpečné toxíny) v prípade dostatku živín premnožia a vytlačia konkurenčné zelené riasy. Zdanlivá čierna je síce v skutočnosti len tmavo-červená (prípade v závislosti od použitých fykobilínov aj tmavo-modro-zelená), avšak je to názorná ukážka snahy prírody využiť dostupné žiarenie čo najefektívnejšie. Pre zaujímavosť si ešte uveďme, že netoxické druhy kyanobaktérií (konkrétne rodu Arthrospira) sú používané ako kvalitný zdroj obživy a do veľkej miery pokrývajú proteínové potreby ľudského organizmu.
Vľavo - "čierny" lišajník. Ide o živý symbiotický organizmus tvorený kyanobaktériou a hubou. Kyanobaktéria využívajúca fykobilíny spôsobuje vďaka čiastočne efektívnejšiemu pohlcovaniu spektra zdanlivo čiernu farbu, v skutočnosti však ide o tmavo modrozelenú farbu, nepohlcuje sa rovnomerne celé spektrum. V strede - kyanobaktérie na spodnej časti kameňa. Vpravo: kyanobaktérie v akváriu. Zdroj: Saratoga Woods and Waterways (vľavo)
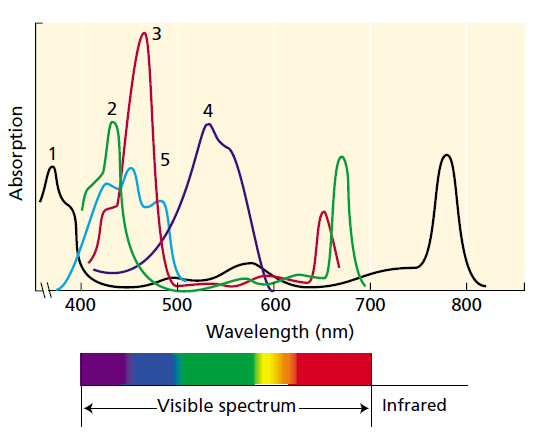
Obr. A-4: Fykoerytrín používaný napr. kyanobaktériami pohlcuje najmä zelenú oblasť spektra. Efektivita pohltenia viditeľného spektra je vyššia než u rastlín, avšak napriek tomu nepokrýva celé spektrum- Používajú ho organizmy, ktoré nepotrebujú vyvíjať molekulárne chladiace mechanizmy (napr. niektoré lišajníky, ale predovšetkým organizmy žijúce vo vodnom prostredí). Na obrázku vidíme absorpčnú schopnosť fykoerytrínu (zodpovedajúcu krivke 4) v porovnaní s najrozšírenejšími fotopigmentmi: 1 - baktériochlorofyl a, 2 - chlorofyl a, 3 - chlorofyl b, 5 - betakarotén. Zdroj: L. Taiz, E. Zeiger: Plant Physiology, Sinauer Associates; 3rd edition (2002), Chapter 7, p. 115.
B. Makroskopická štruktúra rastlín sa vyznačuje vrstvením listov, inými slovami listy rastliny sa prekrývajú a vytvárajú tak mnohovrstvovú absorpčnú plochu. Časť žiarenia (vrátane toho najmenej pohlcovaného zeleného), ktoré nepohltia vyššie položené listy, pohltia listy položené nižšie, a preto v mnohých prostrediach dokonca aj fotóny zelenej časti spektra sú postupne pohltené takmer všetky. To znižuje evolučný tlak na náročný vývoj efektívnejších pigmentov, keďže už aj súčasné rastliny dokážu zelené fotóny pohlcovať pomerne efektívne (hoci nie najefektívnejšie), a kvalitatívne nedostatky na molekulárnej úrovni dokážu čiastočne kompenzovať kvantitou (inými slovami "hrubou silou"), v tomto prípade viacerými vrstvami listov na jednej rastline.
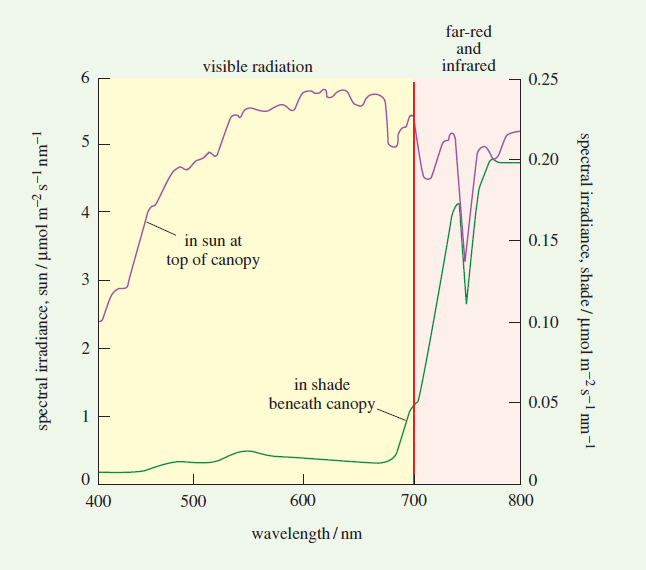 Obr. B-1: Spektrálna intenzita slnečného žiarenia nad (horná krivka) a pod (spodná krivka) klenbou rastlinstva vo vegetáciou husto pokrytom prostredí, napr. dažďovom pralese, v jednotkách relevantných pre chemický proces fotosyntézy. Vidíme, že v takýchto rastlinných spoločenstvách sa aj zelená časť spektra (~500-570 nm) do značnej miery pohltí, i keď neustále tvorí najväčšiu časť nepohltenej časti viditeľného spektra. Graf ilustruje aj snahu rastlín nepohlcovať infračervené fotóny (vo všeobecnosti primárny zdroj prehrievania). Zdroj: I. Ridge: Plants, Oxford University Press, Chapter 02, p. 61.
Obr. B-1: Spektrálna intenzita slnečného žiarenia nad (horná krivka) a pod (spodná krivka) klenbou rastlinstva vo vegetáciou husto pokrytom prostredí, napr. dažďovom pralese, v jednotkách relevantných pre chemický proces fotosyntézy. Vidíme, že v takýchto rastlinných spoločenstvách sa aj zelená časť spektra (~500-570 nm) do značnej miery pohltí, i keď neustále tvorí najväčšiu časť nepohltenej časti viditeľného spektra. Graf ilustruje aj snahu rastlín nepohlcovať infračervené fotóny (vo všeobecnosti primárny zdroj prehrievania). Zdroj: I. Ridge: Plants, Oxford University Press, Chapter 02, p. 61.
C. Nie je vylúčené, že nám vďaka nášmu ohraničenému poznaniu živej prírody uniká jeden či viac relevantných faktorov, ktoré v prírode existujú, a ktoré popri fyzikálno-chemickej efektivite vstupujú do celkového cyklu života rastlín, pričom môžu neefektívnu absorpciu zelených fotónov zvýhodňovať. Je to však pomerne málo pravdepodobné, keďže proces fotosyntézy a energetického života bunky poznáme dostatočne dobre na to, aby sme mohli tvrdiť, že môžu existovať iba faktory brzdiace evolúciu smerom k najefektívnejšej čiernej farbe (čiže vytvárajúce bariéry medzi lokálnymi minimami), nie však faktory, ktoré by spôsobovali, že čierna farba nemôže byť jednou z charakteristík globálneho minima evolučného vývoja. Ani dôležité faktory ako napr. prehrievanie listov (vďaka príliš intenzívnej absorpcii žiarenia) neposkytujú odôvodnenie mrhania zelenou časťou spektra. Prehrievanie totiž spôsobujú najmä vysokoenergetické modré fotóny (vďaka tomu, že spôsobujú nestabilitu chlorofylu – pre podrobnosti viď druhú časť bodu E) a nízkoenergetické infračervené fotóny (pohlcované vďaka ich rezonancii s molekulami vody obsiahnutými v bunkách), nie zelené. Ak by bol teda problémom prebytok či intenzita žiarenia, tak by sa rastliny buď vedeli brániť na makroskopickej úrovni (napr. vhodným tvarom či natáčaním listov v závislosti od dennej doby a sklonu slnečných lúčov, tak ako to robia mnohé rastliny aj dnes), prípadne by sa na mikroskopickej úrovni mali snažiť nepohlcovať predovšetkým modré a infračervené fotóny (a teda by mali byť modré, nie zelené). Keďže však rastliny evidentne modré nie sú, tak dôvodom ignorancie zelenej časti spektra nemôže byť prehrievanie. Dôvodom by mohlo byť nanajvýš pre prípad použitia chlorofylu, ktorý zo svojej definície modrú časť spektra absorbuje síce veľmi dobre, ale práve za cenu toho, že spôsobuje značné prehrievanie – tu sa opäť dostávame k bodu A, keď by vývojová cesta chlorofylu predstavovala iba lokálne minimum evolučného vývoja, a môže byť príčinou, prečo suchozemské rastliny nepoužívajú síce efektívnejšie, ale stále iba s chlorofylom prepojené fykobilíny. V každom prípade, pre vyriešenie problému príliš intenzívneho ožiarenia predovšetkým vlnovými dĺžkami modrej farby si rastliny vyvinuli aj na molekulárnej úrovni sofistikované spôsoby ochrany (nazývané všeobecným pojmom fotoinhibícia). Aj keď ani táto skutočnosť neodôvodňuje mrhanie zelenou časťou spektra, môže nám pomôcť pochopiť, prečo sa zatiaľ nevyvinuli čierne rastliny. Nový fotopigmentový komplex totiž musí zároveň okrem efektivity pri slabom žiarení (napr. v ranných či večerných hodinách, v zime či počas zamračenej oblohy) vyvinúť aj účinnú ochranu pred intenzívnym žiarením (napr. na poludnie, či v prípade vegetácie existujúcej v tieni iných rastlín pri krátkodobých ožiareniach plným denným svetlom). Maximálna efektivita teda musí byť funkčne prepojená s flexibilitou účinnosti fotosyntézy pri rôznych úrovniach intenzity žiarenia, čím sa vývoj nového fotopigmentového systému komplikuje. Možno práve preto je zatiaľ schopnosť fotosyntézy pohlcovať aj zelenú časť spektra (i keď stále nie celé viditeľné spektrum) obmedzená len na niektoré prehrievaniu nevystavené organizmy. V tejto súvislosti je užitočné si uvedomiť aj tú skutočnosť, že pred niekoľkými miliardami rokov v dobe vzniku chlorofylu samotné Slnko žiarilo o niekoľko desiatok percent menej intenzívne než dnes, takže v tej dobe fotoinhibícia nebola natoľko (ak vôbec) potrebná - v minulosti teda ešte viac než dnes nebol dôvod na obmedzenie pohlcovania zelenej časti spektra.
Kľúčovým parametrom nemôže byť ani skutočnosť, že prvé fotosyntetizujúce organizmy sa vyvinuli v morskej vode, v ktorej je spektrum žiarenia odlišné od žiarenia na súši (voda je účinným filtrom červenej časti spektra), pretože to by síce vysvetľovalo citlivosť chlorofylov na modrú časť spektra, avšak nevysvetľuje (naopak, robí ešte viac nelogickou) ich citlivosť na oblasť červenú pri súčasnej ignorancii oblasti zelenej.
V tejto súvislosti je zaujímavé spomenúť aj výnimočnú schopnosť, ktorú si vyvinuli niektoré organizmy, a to nielen žijúce v podmienkach, kde by iné fotosyntetizujúce organizmy neprežili. Predovšetkým vďaka už spomínaným fykobilínom sú totiž schopné flexibilne meniť zloženie fotopigmentov a tým aj celkovú absorpčnú schopnosť svojich fotosystémov. Dôsledkom toho je aj zmena ich farby, čo poskytlo základ pre pomenovanie javu - komplementárna chromatická adaptácia. Túto zmenu dokážu uskutočňovať veľmi rýchlo, nie dlhodobým evolučným vývojom – baktéria si vytvorí najvhodnejšiu kombináciu koncentrácií pigmentov už priamo počas svojho rastu. Možno neprekvapí, že priekopníkom v tejto flexibilnej maximalizácii efektivity fotosyntézy sú práve kyanobaktérie, pôvodcovia chlorofylu.
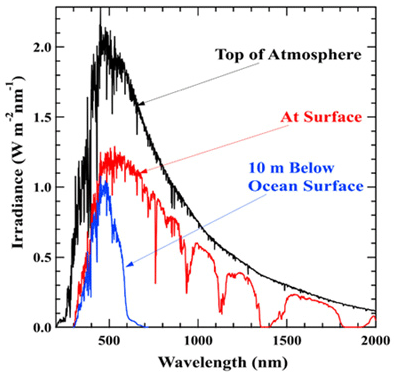
Obr. C-1: Spektrálne rozloženie energetického príkonu žiarenia dopadajúceho na vrchnú časť zemskej atmosféry zo Slnka (čierna krivka), na povrch Zeme po prefiltrovaní atmosférou (červená krivka) a v hĺbke desať metrov pod povrchom morskej hladiny (modrá krivka). Viditeľné žiarenie sa nachádza v oblasti spektra približne od 390 nanometrov (nm) do 750 nm. Vidíme teda, že maximum energie dopadajúcej na povrch Zeme aj oceánov je vo viditeľnej časti spektra (nie je teda žiadnou náhodou, že i ľudský zrak sa evolučne vyvinul na vnímanie práve tejto najintenzívnejšej oblasti), a teda aj fotosyntetizujúce rastliny sa veľmi racionálne sústreďujú vo všeobecnosti na túto oblasť, s výnimkou zelenej časti (cca 500 - 570 nm). Túto výnimku nie je možné vysvetliť ani tým, že sa fotosyntéza a chlorofyl vyvinuli v oceánoch s odfiltrovanou červenou (a vo väčších hĺbkach čiastočne aj zelenou) časťou spektra, keďže červenú časť chlorofyl využíva, a keďže jeden z najprogresívnejších fotosyntetizujúcich organizmov – kyanobaktérie – evolučne vyvinuli už aj zelenú časť spektra absorbujúce do chlorofylových anténnych systémov zakomponovateľné fotopigmenty (fykobilíny). Zdroj: Laboratory for Atmospheric and Space Physics, University of Colorado.
D. Zelená farba rastlín môže byť aktuálnym produktom evolúcie vďaka tomu, že je pravdepodobné, že prvotné fotocitlivé organizmy mali pigmenty absorbujúce predovšetkým zelenú časť spektra (spomínaná skupina fotopigmentov retinalu, ktorá existuje dodnes). Vo vrchnej vrstve vody (na súši život vtedy nejestvoval) teda bolo zelené svetlo značne absorbované množstvom vtedy dominujúcich organizmov skupiny Archaea, a preto sa evolúciou vyvinuli pigmenty absorbujúce zostatkové červené a modré žiarenie, ktoré prenikalo aj do väčších hĺbok, ponechávajúc tým evolučný priestor konkurenčným mikroorganizmom. Tie neskôr vďaka vyššej efektivite nimi vyvinutej konkurenčnej fotosyntézy a meniacim sa globálnym podmienkam na Zemi v ďalšej evolúcii získali prevahu a dnes tvoria väčšinu fotosyntetizujúcich organizmov, avšak doposiaľ nevyvinuli kombináciu do chlorofylového komplexu zapojiteľných fotopigmentov pohlcujúcich rovnomerne celé viditeľné spektrum (čím sa dostávame opäť k bodu A).
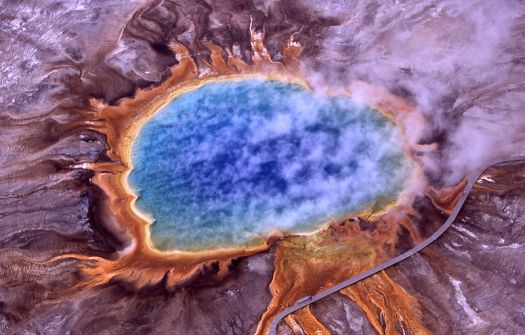 Obr. D-1: Odlišnosti mikroorganizmov patriacich do skupiny (domény) Archaea im umožňujú na rozdiel od väčšiny baktérií a eukaryotických (čiže rastlinných, živočíšnych a iných) buniek žiť v extrémnych podmienkach, napr. sa im výborne darí v slaných prostrediach v Mŕtvom mori či horúcich a kyslých vodách Yellowstonského národného parku (jedno z jeho jazierok s veľkosťou cca 100 metrov sfarbeného archaeami vidíme na obrázku v prirodzených farbách). Ich purpurovo červená farba nie je výsledkom fotosyntézy (a to ani s pomocou karotenoidov či fykobilínov, ktoré u niektorých fotosyntetizujúcich baktérií majú za následok podobnú farbu), ale je výsledkom použitia menej komplexného retinalu a s ním spojeným (nie fotosyntetických, iba fototropných) fotopigmentov, predovšetkým tzv. baktériorodopsínu (ktorý ale nie je využívaný baktériami, jeho názov je historickým prežitkom z doby, keď sa doména Archaea nekorektne zaraďovala medzi baktérie). Zdroj: U.S. National Park Service.
Obr. D-1: Odlišnosti mikroorganizmov patriacich do skupiny (domény) Archaea im umožňujú na rozdiel od väčšiny baktérií a eukaryotických (čiže rastlinných, živočíšnych a iných) buniek žiť v extrémnych podmienkach, napr. sa im výborne darí v slaných prostrediach v Mŕtvom mori či horúcich a kyslých vodách Yellowstonského národného parku (jedno z jeho jazierok s veľkosťou cca 100 metrov sfarbeného archaeami vidíme na obrázku v prirodzených farbách). Ich purpurovo červená farba nie je výsledkom fotosyntézy (a to ani s pomocou karotenoidov či fykobilínov, ktoré u niektorých fotosyntetizujúcich baktérií majú za následok podobnú farbu), ale je výsledkom použitia menej komplexného retinalu a s ním spojeným (nie fotosyntetických, iba fototropných) fotopigmentov, predovšetkým tzv. baktériorodopsínu (ktorý ale nie je využívaný baktériami, jeho názov je historickým prežitkom z doby, keď sa doména Archaea nekorektne zaraďovala medzi baktérie). Zdroj: U.S. National Park Service.
E. Proces fotosyntézy zahŕňa okrem fotopigmentov aj ďalšie proteínové komplexy, z ktorých sa každý zúčastňuje na niektorej fáze procesu. Tieto proteíny/enzýmy tiež majú svoje obmedzenia. Faktorom, ktorý najviac limituje súčasnú fotosyntézu v rastlinách, je zakomponovanie molekuly oxidu uhličitého do organickej molekuly (tzv. fixácia uhlíka, ktorá je súčasťou druhej fázy fotosyntézy nezávislej od svetla), ktoré zabezpečuje enzým nazývaný RuBisCO. Tento veľmi dôležitý proces (vďaka čomu je RuBisCO najrozšírenejším proteínom v prírode, a tvorí 30 až 50 percent všetkých proteínov v listoch rastlín) však uvedený evolučne prastarý enzým realizuje veľmi neefektívne, a to aj v porovnaní s väčšinou iných enzýmov v organizmoch (pre porovnanie, zatiaľ čo mnohé enzýmy dokážu za sekundu uskutočniť stovky až tisícky chemických reakcií, RuBisCO uskutočňuje v priemere iba dve až tri). Ďalšou nevýhodou je schopnosť tohto enzýmu viazať kyslík pri nižšej koncentrácii oxidu uhličitého, čo má za následok až úplné zastavenie fotosyntézy pri podkritickom obsahu CO2 v atmosfére. Je teda logické, že ak máme vylepšiť fotosyntézu ako celok, ďalšie vylepšenie už pomerne efektívneho pohlcovania fotónov prinesie len malý efekt, pokiaľ budú zvyšné súčasti procesu naďalej vysoko neefektívne.
Takáto degradácia prípadných vylepšení len ďalej zníži evolučnú výhodu, ktorú by prípadný vývoj efektívnejšieho fotosystému so sebou priniesol, a teda predĺži čas potrebný na ich realizáciu. Pozemské organizmy sa pokúšajú nahradiť molekulu RuBisCO efektívnejšími alternatívami, avšak úspech bol len čiastočný a žiadna z alternatív zatiaľ nie je plnohodnotnou náhradou.
Obr. E-1: Hlavným limitujúcim faktorom efektívnosti dnešného procesu fotosyntézy nie je absorpčná schopnosť fotopigmentov, ale nízka rýchlosť reakcií enzýmu RuBisCO, ktorý je v porovnaní s ostatnými enzýmami veľmi pomalý. Prírode sa zatiaľ podarilo nájsť len čiastočné náhrady a teda kľúčový faktor pri vylepšení fotosyntézy na ňu ešte čaká predovšetkým v nájdení efektívnejšej alternatívy tohto evolučne veľmi starého a na zemi najrozšírenejšieho enzýmu. Keďže sa jej zatiaľ plne táto úloha nepodarila, nie je prekvapujúce, že menší zdroj neefektivity (nepohlcovanie zelenej časti spektra) zostáva zatiaľ tiež nevyriešený. Zdroj: wikimedia.
Okrem fixácie uhlíka sa pri fotosyntéze vyskytujú aj ďalšie straty energie, ktoré síce nie sú až natoľko závažné ako v prípade RuBisCO, avšak stále prevyšujú straty spôsobené zelenou farbou rastlín – konkrétne napr. straty pri pohlcovaní modrej časti spektra. Pri pohltení týchto vysokoenergetických fotónov je totiž molekula chlorofylu vysoko nestabilná a prebytočnej energie sa rýchlo zbavuje vo forme tepla, pričom si ponechá len energiu ekvivalentnú pohlteniu nízkoenergetického červeného žiarenia. Táto degradácia energie fotónov modrej časti spektra na nižšie energie predstavuje značnú stratu (približne 25 percent celkovej využiteľnej energie viditeľného spektra) a navyše aj problém pri prehrievaní fotosystémov. Fotopigment s vyššou stabilitou po absorbovaní vyššej energie než má chlorofyl by teda bol dvojnásobne prospešný, okrem zvýšenia efektivity fotosyntézy by citeľne znížil prehrievanie rastlín.
Zelená farba rastlín teda nie je zďaleka primárnym zdrojom neefektivity fotosyntézy a je vcelku prirodzené, že mierne dolaďovanie efektivity nebude mať efekt pred tým, než sa zoptimalizujú dôležitejšie nedostatky. Evolúcia však postupuje - optimalizačný proces vidíme napr. aj priamo na našich poliach – druhú (od svetla nezávislú) časť fotosyntézy (tzv. Calvinov cyklus, ktorého detaily boli mimo rámca tohto článku, preto sme sa im nevenovali, a ktorého súčasťou je i fixácia uhlíka) niektoré rastliny čiastočne zdokonalili, vďaka čomu zvýšili efektivitu premeny energie fotónov na chemickú energiu vo vhodných klimatických podmienkach o niekoľko percent. Sú to tzv. „C4 rastliny“, ktoré sa vyvinuli iba pred cca 30 miliónmi rokmi, a medzi ktoré patrí napr. zhruba polovica druhov tráv (vrátane kukurice či cukrovej trstiny). Táto zmena však nezabezpečila, že všetky rastliny dnes používajú zdokonalený C4 cyklus - je to zapríčinené tým, že C4 proces zefektívňuje fotosyntézu iba pri vyšších teplotách, nízkej vlhkosti a nízkom obsahu oxidu uhličitého v atmosfére. C4 rastliny tvoria preto sotva 5 % biomasy Zeme, avšak uskutočňujú až 30 % fixácie uhlíka. V každom prípade, keďže ani C4 rastliny zďaleka nevyriešili neefektivitu enzýmu RuBisCO, tak na vylepšenie absorpcie fotónov (čierna farba by zvýšila efektivitu o niekoľko percent, čiže mnohonásobne menej než sú straty spôsobované enzýmom RuBisCO) si príroda s najväčšou pravdepodobnosťou ešte dlhý čas počká. Je to podobná situácia, ako by sme chceli napr. na predvojnovom automobile znížiť spotrebu paliva – efekt novej aerodynamickej karosérie bude minimálny a nekonkurencieschopný, pokiaľ nedôjde k výmene či podstatnej úprave motora.
F. Na výrobu pigmentov je potrebná (chemická) energia. Keďže zabezpečiť čo najrovnomernejšiu absorpciu celého viditeľného spektra žiarenia znamená vyrábať veľké množstvo fotopigmentov, nie je možné s istotou vylúčiť možnosť, že príroda predsa len našla optimálny kompromis medzi energetickou náročnosťou výroby pigmentov a medzi energetickým ziskom, ktorý by takýto pigment priniesol. Tento argument však nie je vo vedeckej obci všeobecne akceptovaný, predovšetkým z toho dôvodu, že (ako sme už spomenuli) zelená časť spektra prináša na povrch Zeme veľmi významnú časť energie. Je teda nepravdepodobné, že by energetická náročnosť výroby pigmentov bola dôvodom, prečo nevyužiť naplno tento bohatý zdroj energie, keď iné menej energeticky prospešné časti spektra pigmentmi pohlcované sú.
Iba 'akademická' otázka?
Mnohým pri čítaní týchto riadkov pravdepodobne skrsne v hlave myšlienka, že otázka tohto článku je možno síce nie nezaujímavá, ale v podstate zbytočná či príliš akademická, bez praktického uplatnenia. Opak je však pravdou. Efektivita fotosyntézy a z toho plynúca schopnosť živých organizmov pohlcovať isté vlnové dĺžky materskej hviezdy môže byť už v blízkej dobe veľmi relevantná v oblasti hľadania extraterestriálneho života. V posledných dvoch dekádach sú veľkou rýchlosťou objavované planéty mimo našej slnečnej sústavy, momentálne ich poznáme niekoľko stoviek a vďaka novým automatizovaným ďalekohľadom na obežnej dráhe okolo Zeme existuje ďalších niekoľko tisíc kandidátov. Podľa najnovších vedeckých odhadov sa v našej Galaxii nachádza viac planét ako je hviezd, a teda počet prinajmenšom mikroorganizmami obývateľných planét bude rovnako nie zanedbateľný. Zo spektrálnej analýzy žiarenia z týchto planét (ktorú už vedci dokázali uskutočniť, a ktorá bude predovšetkým s nasledujúcou generáciou ďalekohľadov rutinnou súčasťou exoplanetárnej astronómie) môžeme určiť, ktoré vlnové dĺžky žiarenia sú pohlcované, a tým aj určiť príčinu danej situácie, vrátane možnej absorpcie žiarenia tamojším fotosyntetizujúcim životom. Je preto užitočné pochopiť a uvedomiť si vzájomnú súvislosť spektrálneho zloženia žiarenia materských hviezd a rôzne možnosti pigmentov prípadného fotosyntetizujúceho života na ich planetárnych obežniciach či ich mesiacoch.
 Obr. 6: Pri podrobnej analýze spektier extrasolárnych planét ďalekohľadmi v blízkej budúcnosti bude pre hľadanie života dôležité mať na zreteli možnosti fotosyntézy a jej variantov. Zdroj: Discover Magazine.
Obr. 6: Pri podrobnej analýze spektier extrasolárnych planét ďalekohľadmi v blízkej budúcnosti bude pre hľadanie života dôležité mať na zreteli možnosti fotosyntézy a jej variantov. Zdroj: Discover Magazine.
Navyše, otázka z nadpisu tohto článku v sebe skrýva aj priamu podotázku - ako dosiahnuť, aby rastliny boli čierne? Inými slovami, ako vylepšiť fotosyntézu až k jej maximálnej efektivite a tvorbe biologických látok a rastu rastlín? Bližšie porozumenie dôvodom neefektivít nám môže umožniť pomôcť rastlinám vo využití dostupnej energie na ich prirodzené procesy, čo by samozrejme malo význam nielen v poľnohospodárstve pri pestovaní plodín, ale napr. i v (hoci kontroverznom) klimatickom inžinierstve pri snahe o efektívnejšie zabudovanie oxidu uhličitého do biologickej hmoty. Podobné snahy zamerané predovšetkým na (racionálne ako primárny zdroj neefektivity identifikovaný) enzým RuBisCO sú už predmetom vedeckého výskumu, podobne ako možné využitie kyanobaktérií pri produkcii chemickej, resp. elektrickej energie. Teoretická hranica efektivity fotosyntézy je totiž niekoľkonásobne vyššia ako súčasný stav, ku ktorému doposiaľ príroda dospela.
Sumarizácia
Je pravdepodobné, že príčinou absencie čiernej farby pozemských fotosyntetizujúcich organizmov je paralelná existencia viacerých faktorov, ktoré prírodu pri jej ceste k najefektívnejšiemu riešeniu, teda čiernym rastlinám, brzdia. Žiaden z nich však túto najefektívnejšiu formu fotosyntézy neznemožňuje. Na záver si tieto faktory stručne zhrňme a zrekapitulujme obsah a súvislosti predchádzajúcej kapitoly:
(I) Evolúcia doposiaľ neskončila, pričom príroda sa pri optimalizácii procesov z počiatočného stavu aktuálne nachádza v jednom z mnohých lokálnych miním (tak ako sa v nich nachádzala aj v minulosti). Navyše, súčasné riešenie pohlcovania spektra je už relatívne efektívne. Na ďalší posun je potrebná súhra viacerých paralelných zmien parametrov [viď bod II tejto sumarizácie], čo si vyžaduje enormne dlhý čas, ktorý zatiaľ príroda nemala k dispozícii, a možno ho ani mať nebude. Indíciou snahy prírody zvyšovať efektivitu fotosyntézy však sú napr. fykoerytríny, ktoré v novátorských anténnych chlorofylových systémoch dokážu účinne absorbovať aj zelenú časť žiarenia. Napriek tomu, ani oni nezabezpečujú rovnomerné pohlcovanie viditeľného žiarenia v chloroplastoch, a navyše sú zatiaľ obmedzené len na vodné prostredie, kde nie je nutnosť vyvíjať sofistikované molekulárne chladiace mechanizmy [viď bod V]. Druhou indíciou progresu prírody smerom k efektívnejšiemu využitiu dostupného spektra žiarenia je komplementárna chromatická adaptácia (čiže flexibilná maximalizácia fotosyntézy pri rôznych druhoch žiarenia) niektorých kyanobaktérií.
(II) Proces vývoja nového či vylepšeného fotosystému si vyžaduje zabezpečiť nielen jeho funkčné a efektívne zakomponovanie do existujúcich komplexných procesov fotosyntézy, ale musí zabezpečiť i molekulárne fotoinhibičné mechanizmy chrániace fotosystémy pred dočasným prebytkom žiarenia, ktoré sa v prírode vyskytuje. Táto bariéra prispieva k predĺženiu času potrebného na evolučný progres.
(III) Existujúce riešenie rastlín aspoň čiastočne pokrýva i zelenú časť spektra (ktorej pohlcovanie chlorofylom je síce nízke, ale zďaleka nie nulové), čo pri mnohovrstvovej makroskopickej štruktúre väčšiny rastlín pomáha pohltiť väčšinu i tejto oblasti žiarenia (a teda riešiť problém kvantitou namiesto kvality), čo v konečnom dôsledku znižuje evolučný tlak na vývoj nového čierneho fotosystému.
(IV) V komplexnom procese fotosyntézy nie je nízka absorpcia zelenej časti spektra najväčším zdrojom neefektivity. Plnohodnotného vylepšenia sa však doposiaľ nedočkali ani viaceré z týchto niekoľkonásobne viac neefektívnych procesov (najmä proces fixácie uhlíka neefektívnym enzýmom RuBisCO a strata energie pri pohltení fotónov modrej časti spektra). V konečnom dôsledku by teda efekt pohlcovania zelených fotónov nepriniesol výrazné zlepšenie celkovej efektivity, keďže by bol do značnej miery znehodnotený či zredukovaný omnoho väčšími stratami v iných fázach procesu.
(V) Je nepravdepodobné, že by súčasná zelená farba rastlín bola konečným najefektívnejším riešením z dôvodu možného prehrievania rastlín či energetickej náročnosti výroby fotopigmentov, keďže absorpcia zelenej časti spektra nekoliduje s optimalizáciou týchto procesov. Prehrievanie však môže byť dôvodom, prečo je zelená farba dočasne zafixovaná už pomerne dlhú dobu (viď bod II), keďže suchozemské rastliny vďaka neefektivite a prehrievaniu chlorofylu pri pohlcovaní modrej časti spektra musia buď vyvinúť súčasne s efektívnejšími sekundárnymi pigmentmi aj účinné molekulárne ochladzovacie mechanizmy, alebo vyvinúť úplne nový typ základného fotopigmentu namiesto chlorofylu, ktorý by nedegradoval veľkú časť energie modrých fotónov na tepelnú energiu.
(VI) Rovnako je tiež nepravdepodobné, že by zelená farba bola stabilným a najefektívnejším konečným riešením v dôsledku prípadných vede skrytých a doposiaľ neodhalených aktuálne existujúcich faktorov. Už na základe súčasných teoretických a experimentálnych znalostí procesu fotosyntézy totiž môžeme tvrdiť, že existujú iba faktory brzdiace evolúciu smerom k najefektívnejšej čiernej farbe, nie však faktory vylučujúce čiernu farbu (čiže maximálnu absorpciu) ako jednu z charakteristík globálneho minima evolučného vývoja. Pravdepodobnejšie je, že zelená farba je dôsledkom evolučnej histórie a pozostatkom vplyvu faktorov dôležitých v minulosti, napr. možná dominantnosť purpurových fotosyntetizujúcich organizmov v prvopočiatkoch existencie života (ktoré vyfiltrovaním zelenej časti spektra ponechali neskoršie vyvinutým fotosyntetickým organizmom k dispozícii predovšetkým modrú a červenú oblasť) a iné v čase ťažko odhaliteľné predošlé existenčné podmienky.
Vyhliadky do budúcna a záver
Výkon našej materskej hviezdy - Slnka - sa vďaka termonukleárnemu vývoju veľmi pozvoľna zvyšuje (pred štyrmi miliardami rokov bol jej výkon zhruba o 30% nižší než dnes). Vďaka zvetrávaniu kremičitých hornín to bude mať za následok pokles obsahu oxidu uhličitého v atmosfére pod hranicu, pri ktorej sú ešte rastliny schopné uskutočňovať fotosyntézu. Pre C3 rastliny (tvoriace 95 % biomasy dnešných rastlín) tento kritický okamih nastane o približne 600 miliónov rokov, pre modernejšie C4 rastliny o približne jednu miliardu rokov. Dnešné rastliny majú teda pred sebou už iba limitovanú dĺžku existencie (pre presnejšiu predstavu o časových proporciách si pripomeňme, že prvá komplexná rastlinná bunka vznikla pred viac než miliardou rokov a prvé suchozemské rastliny sa objavili asi pred 475 miliónmi rokmi). Avšak i keby sa evolúcii podarilo medzičasom ďalej znížiť minimálnu hranicu obsahu CO2 potrebnú na uskutočňovanie fotosyntézy (napr. nahradením enzýmu RuBisCO), i tak zvyšujúci sa výkon Slnka v závislosti od vývoja zemskej atmosféry (najmä pokračujúcej fixácie dusíka, ktorá znižuje hmotnosť atmosféry a skleníkový efekt) spôsobí s najväčšou pravdepodobnosťou o jednu, najneskôr dve miliardy rokov kompletné odparenie pozemských oceánov (podporené taktiež nevyhnutným pohltením nezanedbateľnej časti hydrosféry do zemského plášťa) a pravdepodobne teda i zánik mnohobunkového života. Navyše, približne o jednu až tri miliardy rokov vďaka pozvoľnému prenosu rotačnej energie Zeme do kinetickej energie obehu Mesiaca okolo Zeme (ktorý sa podľa meraní od nej vzďaľuje o cca 3,8 milimetra ročne) sa Mesiac vzdiali od Zeme natoľko, že prestane stabilizovať zemskú rotačnú os, čo bude mať kataklizmatické následky na životné podmienky (sklon rotačnej osy sa bude chaoticky a drasticky meniť podobne ako je to už dnes v prípade klimaticky nestabilného Marsu). Ďalším negatívnym aspektom bude pokles prirodzenej rádioaktivity vnútri Zeme, ktorý o viac než miliardu rokov môže spôsobiť útlm konvekcie (t.j. cyklického vzostupu teplejšej a poklesu chladnejšej hmoty) v zemskom plášti, teda i kontinentálneho driftu, a následne i sopečnej činnosti, ktorá prestane dopĺňať oxid uhličitý do atmosféry. Rovnako i už najmenej miliardu rokov trvajúce tuhnutie vonkajšieho zemského jadra spôsobí odhadom o 3 - 4 miliardy rokov zánik zemského magnetického poľa, následné zvýšenie intenzity kozmického žiarenia (a teda radiácie na povrchu Zeme) a stratu atmosférickej vody (vplyvom rozpadu molekúl vodnej pary na kyslík a vodík, ktorý unikne do medziplanetárneho priestoru). V každom prípade, ak by sa Zemi podarilo nejakým neočakávaným spôsobom vyhnúť týmto pravdepodobným scenárom (resp. ak by sa ich naplnenie kvôli neznámym či dnes obtiažne kvantifikovateľným faktorom odsunulo do neskorších období), s istotou môžeme povedať, že o približne päť miliárd rokov Slnko vyčerpá zásoby vodíka vo svojom jadre, vďaka čomu vstúpi do ďalšieho štádia svojho vývoja (v astrofyzikálnej terminológii nazývaného červený obor), ktoré bude znamenať definitívnu likvidáciu možnosti povrchového života na Zemi (a pravdepodobne aj Zeme ako planéty).
Nie je teda isté, či evolúcia bude mať vôbec na čiernu farbu rastlín, ktorá je energeticky najefektívnejším spôsobom využitia energie našej materskej hviezdy, dostatok času. Z geologického a evolučného hľadiska už relatívne onedlho bude musieť riešiť otázku prežitia rastlín (a neskôr i fotosyntetizujúcich baktérií) v kriticky nehostinných podmienkach spôsobených našou starnúcou materskou hviezdou, pri ktorom niekoľkopercentné zlepšenie efektivity pohlcovania žiarenia rozhodne nebude patriť medzi primárne faktory. Keďže fotosyntéza existuje približne 3,5 miliardy rokov, tak má pred sebou už iba kratšiu časť svojej existencie, pričom všetky dnešné rastliny s najväčšou pravdepodobnosťou vyhynú už najneskôr o polovicu tejto doby, prenechajúc bremeno vývoja opäť jednobunkovým organizmom, predovšetkým kyanobaktériám.
V každom prípade, napriek tomu, že živá príroda nie vždy vytvára najefektívnejšie procesy či organizmy, tento aspekt je prirodzenou súčasťou jej vývoja, a nemení to vonkoncom nič na obdivuhodnosti toho, čo miliardy rokov trvajúcou evolúciou dokázala a k akým sofistikovaným a obdivuhodne komplexným výtvorom doposiaľ dospela. Je pravdepodobné, že skúmanie spektier exoplanét nám už v blízkej budúcnosti môže prezradiť viac o flexibilných možnostiach prípadnej fotosyntézy vo vesmíre a ďalej prehĺbiť obdivuhodnosť špecifickej formy hmoty zvanej život.
Autor: Juraj Kotulič Bunta, Ph.D
( LinkedIn )
Výber zdrojov a doplňujúcich informácií:
I. Ridge: Plants, Chapter 2 - Photosynthesis, Oxford University Press, 2002
L. Taiz, E. Zeiger:Plant Physiology; Sinauer Associates, 3rd edition (2002)
R.E. Blankenship: Early Evolution of Photosynthesis, Plant Physiology, vol. 154 no. 2, 434-43 (2010)
Wikipedia: Chlorophyll # Why green and not black
MadSci: Why are plants not black?
Wikipedia: Bacteriorhodopsin
Universiteit Leiden: Why are plants not black?
Wikipedia: Rubisco
Ker Than: Early Earth was Purple, Study Suggests (LiveScience)
Wikipedia: Future of the Earth
- Ak chcete pridať komentáre, tak sa musíte prihlásiť
- prečítané 31254x
-


Je ta otazka dobre polozena?
Na zaciatok sa ospravedlnujem, ak som nepostrehol v texte spravnu odpoved, lebo som cital v podstate len uvodnu a zaverecnu cast.
Podla mna otazka Preco rastliny nie su cierne je zavadzajuca. Preco? Pretoze je primarnym poslanim rastliny efektivne konvertovat energiu slnecneho ziarenia? Nie je. Jej prvotnym poslanim, alebo lepsie povedane jej snahou je rozmnozit sa. Ano, zaiste sa najdu aj ine nazory na prvotne poslanie rastliny, ale konverzia energie bude niekde v uzadi celeho zaznamu.
Ak uvazime, ze prvotnym poslanim rastliny je jej rozmnozovanie, potom konverzia energie je len nastroj, nevyhnutna podmienka k rozmnozovaniu.
Akykolvek zivy tvor, nielen rastlina, vsak ziju v rozmanitom prostredi, kde zastupenie tepla, vody a zivin je velmi premenlive. Rastlina preto musi najst svoj modus vivendi. A cierna farba rozhodne nie je tou najvhodnejsou volbou. Cierna farba sice ponuka efektivnu konverziu energie, ale sucasne je aj najnevhodnejsou pre potreby ochrany teploty samotnej rastliny. Cierna rastlina na prudkom slnku prudko spotrebuvava vzacne zasoby vody na svoje chladenie odparovanim. Co vedie k jej uschnutiu a zaniku. A to rozhodne nie je jej ciel, ak chce dokoncit svoj rozmnozovaci proces. Zelena farba zda sa byt kompromisom.
Tot mojich 2 centy do diskusie z hladiska energetickeho, ako botanik a chemik som laik.
Prajem prijemny vecer
miro novak
Dakujem za reakciu a logicky
Dakujem za reakciu a logicky nazor. Mate i nemate pravdu. Mate pravdu v tom, ze aktualny stav prirody je dosledkom kompromisu evolucie medzi fyzikalnou maximalnou efektivitou procesov a podmienkami ci mnohorakymi faktormi, ktore ovplyvnuju parametre zivota - jednym z nich je aj farba. Ten kompromis je vsak len docasny, a prave o tom clanok pojednava - najefektivnejsim stavom je maximalne vyuzitie energie, teda cierna farba, ten vsak moze (hoci vdaka prekazkam i nemusi) nastat az v buducnosti. Nemate vsak pravdu v tom, ze by bolo dovodom prehrievanie. Otazka prehrievania rastlin je skutocne jednou z primarnych myslienok, ktore sa objavia ako mozny dovod pre zelenu a nie ciernu farbu rastlin. Avsak nie je to tak - v clanku je tejto teme venovany odstavec C v sekcii 'Preco ziva priroda nie je maximalne efektivna'. Strucne zhrnute - prehrievanie nemoze byt dovodom zelenej farby rastlin, pretoze pohlcovanie zelenych fotonov sposobuje vyrazne mensie prehrievanie rastlin ako pohlcovanie modrych fotonov - to z toho dovodu, ze - zjednodusene povedane - pri pohlteni modreho fotonu vsetka jeho energia, ktora prevysuje energiu cerveneho fotonu, sa v rastline premeni na odpadovu tepelnu energiu. Jeden pohlteny modry foton teda poskytne rastline len tolko energie vyuzitelnej na chemicke procesy, ako jeden cerveny foton - a cely zvysok rastlinu iba nechcene zahrieva. Je to dane nedokonalostou chlorofylu, ktory nedokaze energiu excitacie z modrych fotonov vyuzit na chemicko-fyzikalne deje, musi sa jej zbavit. Preto pohlcovanie modrych fotonov chlorofylom je velmi stratove a je hlavnym zdrojom prehrievania rastlin. Ak by teda rastliny mali problem prave s prehrievanim, boli by modre - pretoze by modre fotony odrazali. Zjavne nie su, a preto prehrievanie nemoze byt dovodom zelenej farby rastlin. Zelene fotony su menej energeticke ako modre, a teda ak by si mala rastlina vybrat, radsej by mala pohlcovat co najviac zelenych fotonov, a co najmenej modrych. Evolucia vsak mala zviazane ruky - chlorofyl bohuzial pohlcuje modre fotony velmi dobre (a potom sa molekuly mozu prehrievat). Takze prehrievanie je relevantnym faktorom, avsak iba pre nedokolany chlorofyl, ktory ma problem s modrymi fotonmi. Z globalneho hladiska je teda Vami spominany kompromis iba docasnym riesenim prirody, ktora je blokovana nedostatkami chlorofylu - je teda v lokalnom minime vyvoja spominaneho v clanku, nie v globalnom minime, ktorym je - cierna farba. Ak priroda vyvinie novu molekulu, ktora by nemala problem s modrymi fotonmi, a ak by vyvinula este i novy fotopigment (ak by na to nepostacovala i tato samotna nova molekula nahradzajuca chlorofyl) pokryvajuci oblast, ktora je malo pokryta i fykobilinmi, moze byt rastlina cierna bez obav z prehrievania, ktore v sucasnosti vyplyva hlavne - nechcem sa opakovat - z velmi neefektivneho vyuzitia energie fotonov modrej casti spektra. Zelena farba je teda docasna a vyplyva z nedokonalosti biochemickych procesov.
It is by logic that we prove, but by intuition that we discover. [J. H. Poincaré, mathematician (1854-1912)]
a este by som dodatok k
a este by som dodatok k primarnemu cielu rastlin. Tym je - uplne suhlasim - snaha rozmnozit sa a presadit svoje geny. K tomu vsak potrebuju schopnost co najefektivnejsie produkovat biologicku hmotu - potrebuju napr. v pralese prerast konkurentov, aby sa dostali k svetlu, potrebuju (aspon cast vyssich rastlin) vyprodukovat dostatok kvetov pre opelujuci hmyz, vyprodukovat dostatok energiu obsahujucich plodov (obsahujucich cukor) aby ich semena rozniesli zivocichy, dostatok samotnych opat energeticky na vyrobu narocnych semien ci vytrusov na rozmnozenie, ci v nehostinnch podmienkach potrebuju co najefektivnejsie vyuzit nedostatok zdrojov a energie na samotne prezitie. Vo vsetkych pripadoch je velmi vyznamnym faktorom schopnost premienat energiu fotonov na chemicku energiu, pretoze ta im umoznuje okrem ineho zabudovat anorganicky oxid uhlixity do svojich organickych stuktur, cize produkovat biologicku hmotu z hmoty nezivej, ktora rastlinam umozni prave napr. prerast konkurentov ci prezit na miestach, kde to ine organizmy uz nedokazu. Kto bude mat tuto schopnost efektivnejsiu, bude mat evolucnu vyhodu. Ukazuju to napr. v clanku zmienovane (hoci medzi rastliny samozrejme nepatriace) kyanobakterie, ktore ziju i na miestach, kde ine organizmy uz nedokazu prezit, a dokazu to vdaka viacerym evolucnym vyhodam, pricom jednou z nich je prave schopnost pohlcovat zelenu cast spektra (hoci stale nie su cierne, su ovela tmavsie nez bezne rastliny). Dokonca - ako sa v clanku spomina - napr. s pribudajucou hlbkou v mori menia farbu tak, aby co najefektivnejsie pohltili meniace sa spektralne zlozenie slnecneho ziarenia prenikajuceho do vacsich hlbok. Kyanobakterie teda uz ukazuju, ze priroda sa naozaj o co najvacsiu efektivitu premeny slnecnej energie snazi, pretoze efektivnejsie vyuzitie zdroja energie moze byt evolucnou vyhodou. Ma vsak k dispozici len obmedzeny biochemicky arzenal, ktory ju zatial drzi pri nie najefektivnejsej zelenej farbe. Ak by boli rastliny cierne, produkovali by biohmotu efektivnejsie, a rastli a rozmnozovali by sa teda rychlejsie.
It is by logic that we prove, but by intuition that we discover. [J. H. Poincaré, mathematician (1854-1912)]